
فهرست مطالب
مولکولهای زیستی(پروتئین، RNA، DNA ) با گذشت زمان تغییر میکنند. سرعت و الگوی تغییرات این مولکولها، به دانشمندان کمک میکند تا فرآیندهای دگرگشتی را بهتر درک کنند. دگرگشت مولکولی به مطالعهی اين تغييرات ميپردازد.
دگرگشت مولکولي به چه معناست؟
مولکولهاي زيستي در طول زمان بدون تغيير نيستند. پروتئينDNA, RNA , دچار تغيير و دگرگشت ميشوند. دگرگشت مولکولي، شاخهاي از زيستشناسي دگرگشتي است که به تغييرات دگرگشتي در سطح اين مولکولها ميپردازد. برخي مولکولها بهسرعت و برخی دیگر بسیار آرام، دگرگشت مییابند. این تغییرات بهدلیل برهمکنش دو فرآیند پایهاي دگرگشتي رخ ميدهد. اين دو فرآيند عبارتاند از جهش و تثبيت (fixation).
جهشها به دو نوع پايه تقسيم ميشوند. جهشهای ارثی و جهشهای سوماتیک(پیکری). جهشهاي ارثي در سلولهاي زاياي جنسي رخ ميدهند و به ژنوم نسلهای بعدی منتقل میشوند. جهشهای سوماتیک میتوانند در هنگام رشد و تکوین عادی صورت بگیرند. برای مثال این جهش ممکن است در حین تقسیم سلولی رخ دهد. در صورتی که جهش سوماتیک فرآیندهای سلولی خاصی را هدف قرار دهد، میتواند منجر به رشد سلولی کنترل نشده و سرطان شود. تمرکز دگرگشت مولکولی، بر روی تغییرات ژنتیکی ارثی است.
جهشهاي ارثي عمدتاً هنگام همانندسازي DNA براي توليد گامت رخ ميدهند. اين جهشها باعث ارائهی نسخههای ژنتیکی جدیدی به جمعیت میشوند. برای گیاهان و جانوران مولکول ژنتیکی موردنظر DNA است. برای برخی ویروسها مولکول وراثتی RNA است. بخشهایی از ژنوم DNA يا RNA توسط سلول به پروتئين ترجمه ميشوند. بنابراين برخي تغييرات ژنومي منجر به تغيير در توالی پروتئينهاي کدشونده ميشوند. به اين صورت دگرگشت مولکولي در سطح آمينواسيد رخ ميدهد.
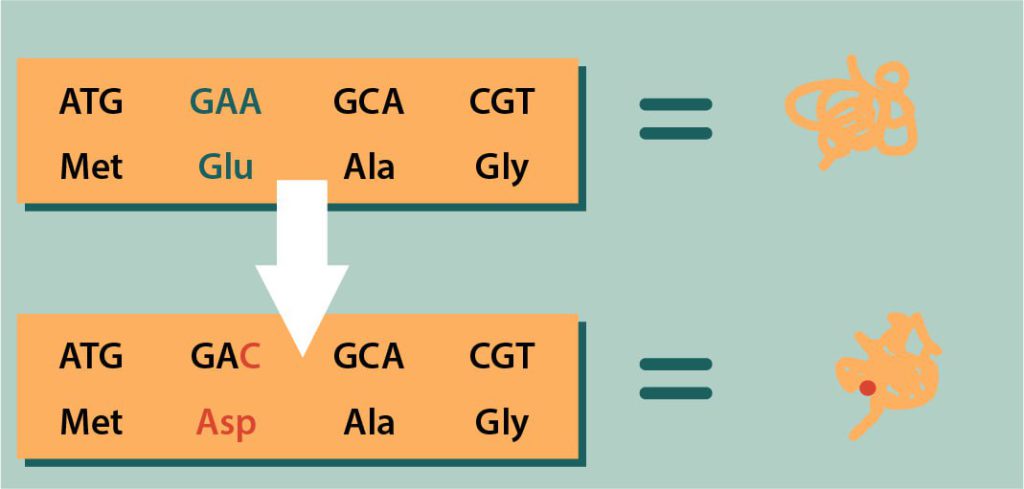
در اينجا تغيير در يک بازآلي منجر به تغيير در توالي آمينواسيدي و توليد پروتئيني جديد شده است.
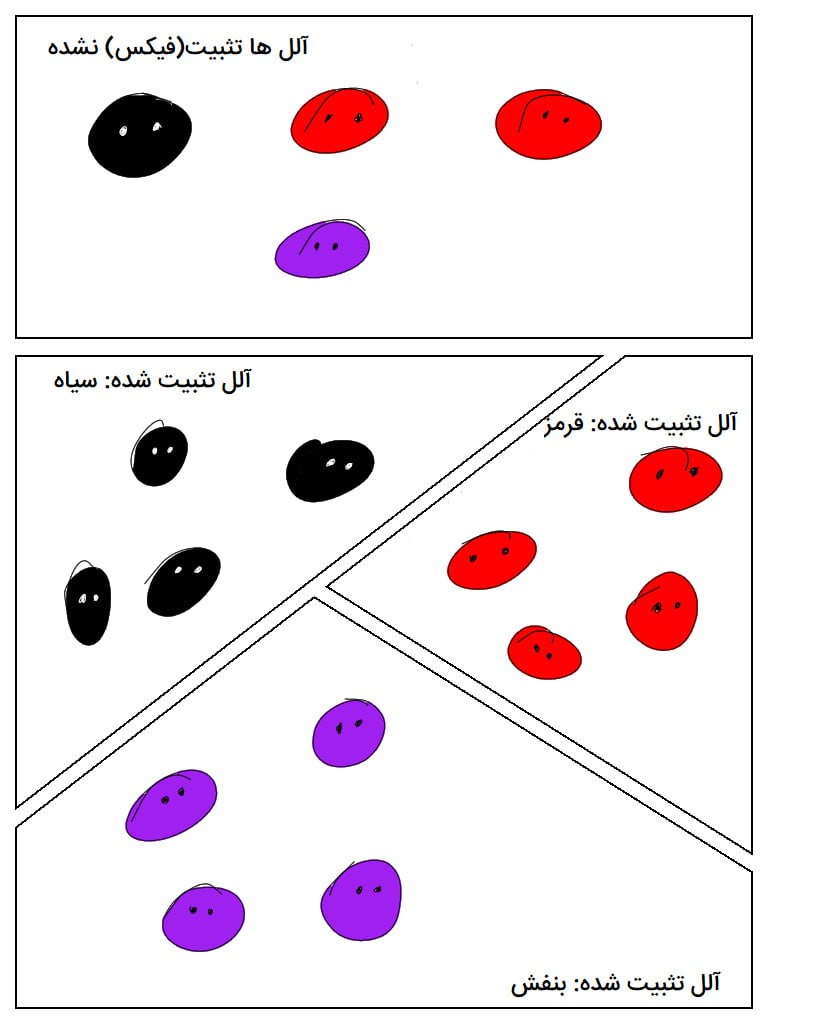
دومين فرآيند دگرگشت مولکولي، تثبيت يا fixation است. تثبيت مرحلهاي جمعيتي است که منجر به تغييرات دگرگشتي در سطح مولکول ميشود. اغلب جهشهای جدید بهدلیل رانش ژنتیکی(تصادفی) و یا بهدلیل مضر بودن از جمعیت حذف میشوند. برخی جهشها به صورت تصادفی و یا بهدليل افزايش موفقيت توليدمثلي در جمعيت باقي ميمانند. رانش ژنتيکي در جمعيتهاي کوچک ميتواند باعث تغييرات سريعي شود ولي در جمعيتهاي بزرگتر اثر کمتري دارد. همچنين نتيجهی رانش به تنهایی، حذف یا تثبیت هر جهش جدید است. به زبان یگر هر جهش در نهایت فراوانی 0 و یا 100 درصدی پیدا میکند. تثبیت به این معناست که جهش جدید جایگزین تمام نمونههای کنونی در ژنوم یک جمعیت شود. تثبیت میتواند بهواسطهي انتخاب طبيعي نيز رخ دهد. انتخاب طبيعي به طور مستقيم باعث افزايش فراواني نسخههايي ميشود که منجر به بقا و موفقيت توليدمثلي ميشوند. اين فرآيند انتخاب مثبت يا به زبان سادهتر سازگاري ناميده ميشود. جهشهای مضر و آنهایی که موفقیت تولیدمثلی را کاهش میدهند عموماً تثبیت نمیشوند.
ساعت مولکولی
در چهل سال گذشته، زيستشناسان دگرگشتی در حال بررسی احتمال بودند که برخی رویدادهای دگرگشتی به شکل ساعتی رخ میدهند. جهشها ممکن است طی میلیونها سال بر روي مناطق معيني از DNA، با سرعتي قابل اطمينان رخ دهند. براي مثال، ژنی که پروتئین آلفا-گلوبین (یکی از اجزای هموگلوبین) را کد می کند، تغییرات پایه را با سرعت 0.56 تغییر در هر جفت باز در هر میلیارد سال تجربه میکند. اگر این سرعت قابل اعتماد باشد، این ژن می تواند به عنوان یک ساعت مولکولی استفاده شود. از ساعت مولکولی میتوان بهعنوان یک ابزار قدرتمند برای تخمین زمان جدایی دودمانها استفاده کرد. برای مثال، تصور کنید بخشی از DNA در دو گونه وجود دارد واین دو بخش در چهار باز آلی با یکدیگر تفاوت دارند. همچنین میدانیم که این بخش از DNA با سرعت تقریبی یک باز در هر ۲۵میلیون سال، تغییر میکند. میتوان نتیجه گرفت که تفاوت بین دو DNA طی ۱۰۰میلیون سال دگرگشت صورت گرفته؛ همچنین نزدیکترین جد مشترک این دو گونه ۵۰میلیون سال پیش میزیسته است. از این تکنیک برای بررسی مسائل مهمی مانند منشاء انسان امروزی، زمان جدایی انسان از شامپانزهها و همچنین زمان انفجار کامبرین استفاده میشود.
نظریهی خنثی
اثار انتخاب طبيعي در همه جا ديده ميشود. اغلب ارگانيسمها به خوبی با محیط زیست خود سازگار شدهاند. با این حال در تمام جمعیتها نسخههاي ژنتيکي ديده ميشود که ممکن است اثري بر روي شايستگي ارگانيسم نگذارند. در دگرگشت مولکولي نظريهی خنثی را مطرح میکند. طبق این نظریه، بسیاری از نسخههای ژنتیکی در جمعیت اثر جهش و رانش ژنتیکی هستند و انتخاب طبیعی در وجود آنها نقشی نداشته است. براساس این نظریه، ممکن است در یک جمعیت آللهاي متفاوتي براي يک ژن وجود داشته باشد و تمامی این آللها به یک اندازه کاربردی باشند. ارگانیسم هر کدام از این آللها را داشته باشد، تفاوتی در میزان سازگاریش ایجاد نمیشود. در واقع این تنوع خنثی است.
نکتهی قابل توجه این است که نظریهي خنثي، جايگزين نظريهي دگرگشت نيست، بلکه شرح بخش کوچکي از نظريهی دگرگشت است. تمام شواهد یافت شده از طریق بررسی نظریهی خنثی، از نظریهي وگرگشت نيز حمايت ميکنند.
دگرگشت ميکروارگانيسمها
دگرگشت میکروبی (microbial evolution)به تغییرات ژنتیکی که در میکروارگانیسمها ایجاد و حفظ میشوند اشاره دارد. برخی از این تغییرات میکروبی در اثر انتخاب بهوجود آمدهاند. بهترين مثال براي اين مسئله، تغييرات متنوعی است که در حضور آنتيبیوتیکها براي باکتريها رخ میدهد. این تغییرات منجر به افزایش مقاومت یا مقاومت کامل باکتری نسبت به کشندگی یک یا چند نوع آنتیبیوتیک میشوند. باقی تغییرات میتوانند در عدم حضور نیروهای انتخابی، به طور تصادفی رخ دهند. این تغییرات اغلب بهدليل تغيير در تک نوکلئوتيدها هستند و ممکن است باعث برتري ميکروارگانيسم نسبت به ميکروارگانيسمهاي بدون تغيير شوند. سپس اين صفت ممتاز حفظ و به نسلهای آینده انتقال داده میشود.
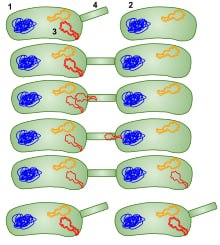
انتقال ژن بين باکتريها، ميتواند بين گونههای غیرمرتبط نیز رخ دهد. این انتقال، انتقال افقی نام دارد و یک نوع مهم از دگرگشت میکروارگانیسمها در طبیعت است. انتقال افقی ژن میتواند در بیماریهاي عفوني نقش داشته باشد؛ براي مثال ميتواند باعث فراگيري ژن مرتبط با مقاومت آنتيبيوتيکي شود.
دگرگشت در میکروارگانیسمها میتواند در طی چند ساعت رخ دهد؛ چراکه برخی باکتریها در شرایط ایدهآل میتوانند هر بیست دقیقه یک بار تقسیم شوند. باکتری حامل ژن تغییریافته و دارای برتری میتواند در عرض بیست و چهار ساعت، هزاران باکتری حامل این ژن را بهوجود آورد. هر یک از باکتریهای ایجاد شده نیز، تا روز بعد هزاران باکتری حامل این ژن تولید میکنند. به این ترتیب جهش به سرعت در جمعیت باکتریایی گسترش میابد. به علاوه، این صفت میتواند به باکتریهای غیرمرتبط نیز منتقل شود. بنابراین جمعیتهای باکتریای دیگر نیز حامل این ژن خواهند شد.
ترسيم تبارزايي يا فيلوژنتيک روابط ميان ميکروارگانيسمها بسيار دشوار است. زيستشناسی مولکولی در مشخص کردن روابط ژنتیکی بین این ارگانیسمها و رسم درخت فیلوژنتیک، بسیار کاربردی است.
دگرگشت پروکاريوتها
قدیمیترین فسیلهای شناخته شده حدود 3.5 میلیارد سال قدمت دارند و متعلق به استروماتولیتها هستند. استروماتولیتها لایههای سنگی هستند که از اتصال تعداد زیادی لابهی باکتریایی و رسوبی به یکدیگر تشکیل شدهاند. فرضیهای وجو دارد که میگوید جانداران تک سلولی از این زمان نیز زودتر منشاء یافتهاند. بر این اساس حیات از حدود 3.9 میلیارد سال پیش شکل گرفته است و پروکاریوتهای اولیه اولین ساکنان زمین بودهاند.

بیشتر اکسیژن اتمسفری، منشاء زیستی دارد و از شکستن آب در فتوسنتز بهوجود آمده است. زمانی که اولین فتوسنتزکنندگان دگرگشت یافتند، اکسیژن آزاد شده احتمالا در آب اطراف حل شد. با افزایش غلطت آن، اکسیژن آزاد با آهن محلول واکنش داد و لایههای رسوبی را بهوجود آورد. پس از آنکه غلظت اکسیژن آزاد در دریاها به حد اشباع رسید، اکسیژن شروع به خروج از آبها کرد و وارد اتمسفر شد. اثر این اکسیژن آزاد بهصورت زنگزدگی سنگهای غنی از آهن، در سطح زمین دیده میشود. این فرآیند از 2.7 ميليارد سال پیش آغاز شده است و نشان میدهد باکتریهای فتوسنتزی آزادکنندهی اکسیژن قبل از 2.7 ميليارد پیش بهوجود آمدهاند. این انقلاب اکسیژنی تاثیر عمیقی بر حیات داشت. غلظت بالای این اکسیژن احتمالا باعث نابودی بسیاری از پروکاریوتها شد. برخی از بازماندگان در زیستگاههای بیهوازی حضور داشتند. در دیگر بازماندگان روشهایی مانند تنفس سلولی، برای سازگاری با اتمسفر ظاهر شدند.
دگرگشت يوکاريوتها
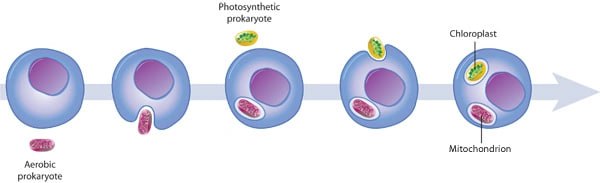
قدیمیترین فسیلهایی که بیشتر محققین بر سر یوکاریوت بودن آنها توافق دارند، سنی حدود 2.1 میلیار سال دارند. چگونه سازمان پیچیدهی سلول یوکاریوتی، از وضعیت سادهی پروکاریوتی بهوجود آمد؟ احتمالاً فرآیندی بهنام درونهمزیستی باعث بهوجود آمدن میتوکندریها و پلاستیدها شده است. طبق تئوری درونهمزیستی میتوکندریها و پلاستیدها در ابتدا پروکاریوتهای کوچکتری بودند که درون سلولهای بزرگتر زندگی میکردند. این اجداد پروکاریوتی احتمالاً به صورت انگلهای درونی و یا شکارهای غیرقابل گوارش، وارد سلول میزبان شدند. اگرچه این فرآیند غیرممکن به نظر میرسد، ولي دانشمندان مواردی از درونهمزیستی را که منجر به ارتباطی سودمند شدند، را مشاهده کردهاند. شروع این ارتباط به هر نحوی بوده باشد، در نهایت به یک همزیستی مفید منجر شده است. به دلیل اینکه پلاستیدها در تمام یوکاریوتها دیده نمیشود، انتظار میرود میتوکندری پیش از پلاستیدها دگرگشت یافته باشد.
در ادامه تعدادی از شواهد تاییدکنندهی این تئوری را بیان میکنیم:
- همانندسازی در میتوکندری و پلاستیدها از طریق یک فرآیند شکافتی، مشابه تقسیم دوتایی باکتری رخ میدهد.
- این اندامکها حاوی DNA حلقوی باکتریایی و فاقد هیستون هستند.
- این اندامکها حاوی RNAهای ناقل، ریبوزومها و دیگر مولکولهای موردنیاز برای رونویسی و ترجمهی DNA به پروتئین هستند.
- غشای داخلی این اندامکها، دارای سیستمهای انتقالی و آنزیمهایی است که مشابه آنها در سلولهای پروکاریوتی زنده یافت میشود.
پس از ظهور اولین یوکاریوتها، طیف گستردهای از تکسلولیهای متنوع به وجود آمد و این تنوع تا به امروز ادامه دارد. برخی یوکاریوتهای تک سلولی ، اشکال پرسلولی را به وجود آوردند. جلبکها، گیاهان، قارچها و جانوران از نسل این یوکاریوتهای پرسلولی هستند. محققان با مقایسهی توالیهای DNA، زمان حضور نیای مشترک یوکاریوتهای پرسلولی را ۱.۵ میلیارد سال قبل برآورد کردهاند. قدیمیترین فسیلهای یافت شده از یوکاریوتهای پرسلولی نیز، متعلق به جلبکهایی هستند که ۱.۲ میلیارد سال پیش میزیستند.
نویسنده: مبینا جاویدی
منابع:
- بیولوژی کمپل؛ ترجمه خانهی زیستشناسي
- https://evolution.berkeley.edu/molecular-clocks/
- https://evolution.berkeley.edu/misconceptions-about-natural-selection-and-adaptation/the-neutral-theory/
- https://edisciplinas.usp.br/pluginfile.php/4481131/mod_resource/content/1/Princeton%20Encyclopedia%20verbete%20de%20evolu%C3%A7%C3%A3o%20molecular.pdf
