
فهرست مطالب
فرگشت مولکولی
مولفان: سعید زارع ۱ ، حسنی ده بزرگی ۲
1-دانشگاه پیام نور ، واحد شیراز ،مرکز ، گروه ریاضیات و کاربرد ها ، شیراز ، ایران
2-دانشگاه دولتی شيراز (دانشکده علوم ) ،گروه زیست جانوری ، شیراز ، ایران
مؤلف رابط : سعید زارع
خلاصه
تکامل مولکولي یا فرگشت ملکولی بخشی از فرایند فرگشت است که در مقیاس دی. ان. ای، آر. ان. ای و پروتئین روی میدهد. در سالهای حدود ۱۹۶۰ میلادی، فرگشت مولکولی به عنوان یک رشته علمی در بین محققان زیستشناسی مولکولی، زیستشناسی فرگشتی و ژنتیک جمعیت، به دنبال درک ساختار و عملکرد اسیدهای نوکلئیک و پروتئینها گسترش پیدا کرد..
مقدمه
فرگشت (به معنای دگرگون شدن و تغییر شکل دادن) بهمعنی دگرگونی در خصوصیتهای وراثتی جانداران در طول زمان است.مجموعه ساز و کارهایی که به سبب ان موجود ی به موجود دیگر تغییر پیدا میکند این تغییرات توسط عواملی همچون انتخاب طبیعی و انتخاب مصنوعی هدایت میشوند این دو عامل با تغییر بر روی ساختار ژن زمینه ساز تغییر در نسل های بعدی و در نتیجه وقوع فرگشت میشوند..
مولکول اساس همه زندگی است و ما میخواهيم بفهمیم که چرا این ملکولها در حال تکامل هستند.
اغلب ما تعاملات میان بخشهای مختلف یک پروتیین را نادیده میگیریم، اما میدانیم که تغییرات در یک بخش از پروتئین تحتتاثیر تغییرات بعدی در دیگر بخشها قرار میگیرد. مشخص مي شود که چرا این مساله برای درک این که چرا این ملکولها به روشی که آنها انجام میدهند تکامل پیدا میکنند مهم است.
پروتیینها به طور مداوم به عنوان جهشهای ثابت تغییر مییابند یا بسته به ساختار پروتیین، عملکرد و پایداری حذف میشوند. این امر بستگی به برهمکنشهای آمینو اسید در سراسر پروتئین دارد که باعث تحول در یک مکان میشوند تا شانس تکامل در سایتهای دیگر را تغییر دهند. دانشمندان کشف کردند که میتوانند میزان تکامل پروتیین را براساس خواص biochemical پیشبینی کنند.
برای سالها، محققان بر ضد مشکلات با مدلهای استاندارد تکامل مولکولی که در مطالعه روابط تکاملی بین گونهها مورد استفاده قرار میگیرند، شرکت کردهاند. این مساله منجر به مشکلاتی در بازسازی رویدادهای مهم تکاملی در ارگانیسمهای اجدادی شد.
این الگوهای همگرایی مولکولی، به طور منظم در طول زمان تکاملی به روشهایی که نشاندهنده محدودیت مستمر در بخشهای مختلف پروتیینها است، یافت شدند. این مساله حول این ایده عادی میچرخد که آمینو اسیدها به نیازهای بقیه پروتئین سازگار خواهند شد. اما ما نمیتوانستیم دقیقا توضیح دهیم که چرا این اتفاق افتاد، یا اینکه آیا هیچ نظمی در این روند وجود داشتهاست ما زمانی که این سیستم در چارچوب مکانیک آماری قرار گرفت، بزرگی اسید آمینه به عنوان مرکزی برای درک میزان واگرایی تکاملی مشاهده شد.
تکامل سلولی
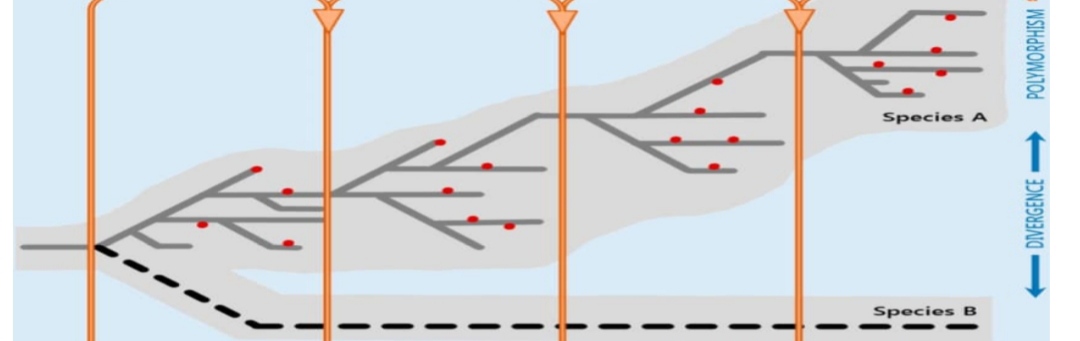
مقایسه مولکول ها یکی از بهترین روش های مطالعه فرگشت است. با پایان یافتن پروژه ژنوم انسان و مقایسه آن با پروژه ژنوم شامپازه مشاهده شد که این دو ژنوم تنها ۱. ۶٪ اختلاف دارند. با پیشرفت روش های استخراج ژن دانشمندان توانستند در یک پروژه حیرت انگیز، ژنهای یک فسیل ۳۲ هزار ساله انسان نئاندرتال (گونه ای منقرض شده از جنس انسان) را استخراج کرده و رابطه مولکولی این فسیل را با انسانهای ساپینس(Homo sapiens) (گونه خود ما و تنها گونه باقی مانده از جنس انسان) را مشخص نمایند. وارسی مولکولی فرگشت تنها به مقایسه کدهای مولکول ها محدود نمی شود، بلکه در سطحی بالاتر به ساختار سه بعدی و بخش عمل کننده ی مولکول ها، خواص بیوفیزیکی آنها و سیستم های شبیهسازی فرگشت مولکولی نیز توجه می شود. از سوی دیگر، مسائلی که در بخش آناتومی مقایسهای ذکر شد در بخش مولکولی هم وجود دارد: همسانی، همساختی، همسویی، ساختارهای ناقص و ساختارهای باقیمانده و بی فاید محققان گفتند که قدرت انتخاب در تکامل پروتیین توسط آنتروپی سلسله تاخوردگی متعادل میشود، تعداد of که پروتیین را با درجه خاصی از ثبات فراهم میکنند.
مرکز سرطان MD اندرسون گزارش می دهد که آنها با موفقیت عنصر تنظیم کننده ژن Prx1 موش را با منطقه تنظیم کننده ژن Prx1 از یک خفاش تغییر داده اند – و اگرچه این دو گونه با میلیون ها سال تکامل از هم جدا شده اند. – موشهای تراریخته بهطور غیرعادی اندامهای بلندی را نشان دادند. در حالی که طول اندام جلویی تنها یکی از چندین تغییر مورفولوژیکی کلیدی است که در طول تکامل بال خفاش رخ داده است، این یافته بیسابقه نشان میدهد که تکامل را میتوان با تغییر در الگوهای بیان ژن هدایت کرد. نه صرفاً با تغییر در ژنهاي خودشان. Prx1 یک فاکتور رونویسی همودومین با جعبه جفتی است که نقش ثابتی در رشد استخوان اندام دارد. دکتر بهرینگر و همکارانش یک دامنه تقویتکننده Prx1 حفاظتشده را شناسایی کردند که بیان Prx1 را در اندام جلویی در حال رشد تنظیم میکند.
دکتر بهرینگر و همکارانش برای مطالعه سهم تکاملی تقویتکننده Prx1 در تفاوتهای مورفولوژیکی بین اندام جلویی خفاش و موش، تقویتکننده درونزای Prx1 موش را با خفاش جایگزین کردند. موش های تراریخته سطوح بیان بالاتری از Prx1 را در پریکندریوم، افزایش تکثیر سلول های غضروفی و در نهایت اندام جلویی بلندتر نشان دادند.[1]
دکتر بهرینگر اهمیت یافته خود را چنین توصیف می کند: «داروین پیشنهاد کرد که” تغییرات جزئی پی در پی «در نهایت منجر به تکامل مورفولوژی های متنوع اندام، مانند دست، بال یا باله می شود. تغییر ژنتیکی که ما در موش ها مهندسی کردیم ممکن است باشد. یکی از آن “اصلاحات جزئی» برای تکامل بال پستانداران.»



تاریخچه تفکر فرگشتی
باور به فرگشت و این که انسانها و دیگر جانداران زنده از راههای طبیعی و دگرگون شدن یک ساختار به ساختاری دیگر پدید آمدهاند ریشه در میان یونانیان باستان، رومیان، چینیان، و علوم اسلامی سدههای میانی دارد. با اغاز طبقهبندی علمی جانداران ، اندیشه زیستی در غرب تحت تأثیر دو باور متضاد قرار گرفت: یکی اعتقاد به این بود که هر جانور دارای خصیصههای ذاتی نامتغیر است که برآمده از مکتب ارسطو و همسو با الهیات طبیعی بود؛ و دیگری برآمده از رویکردی متضاد با اندیشه ارسطویی در دانش نوین بود. در آغاز سده ۱۹ میلادی، ژان–باتیست لامارک نگره خود درباره تراجهش گونهها را مطرح کرد، که نخستین نگره از این نوع درباره فرگشت بهشمار میرفت.
در ۱۸۵۸، چارلز داروین و آلفرد راسل والاس، به صورت مشترک اقدام به چاپ نگرهی فرگشتی تازه کردند. اگرچه تا پیش از ارائه نظریه فرگشتی داروین دیدگاه فرگشتگرایانه در جامعه علمی جای پا باز کرده بود، اما نظریه او به آن دلیل انقلابی تلقی میشود که توانست سازوکاری از چگونگی پدید آمدن تفاوتها در گونههای مختلف را توضیح دهد؛ چیزی که امروزه با مفهوم انتخاب طبیعی میشناسیم. نظریه او همراه بود با شواهد بیشماری از شاخههای گسترده علمی همچون دامداری، جغرافیای زیستی، زمینشناسی، ریختشناسی، و رویانشناسی.

بحث بر روی کار داروین باعث پذیرش سریع مفهوم کلی فرگشت شد، ولی سازوکار مشخصی که تحت عنوان انتخاب طبیعی او پیشنهاد داد، به صورت گسترده مورد استقبال قرار نگرفت، تا پیش از آن هنگام، بیشتر زیستشناسان ادعا میکردند عوامل دیگری مسئول فرگشت هستند. جایگزینهای انتخاب طبیعی که در طی افول داروینیسم پیشنهاد شدند شامل به ارث بردن ویژگیهای اکتسابی میل ذاتی به تغییر و جهشهای آنی و بزرگ بودند. سنتز انتخاب طبیعی توسط ژنتیک مندلی در طی دهههای ۱۹۲۰ و ۱۹۳۰ باعث بنیان نهادن شاخه جدید ژنتیک جمعیت شد. سنتز فرگشتی نوین در پی استقرار زیستشناسی فرگشتی، مطالعات جهش و تنوع در جمعیتهای طبیعی، در ترکیب با جغرافیای زیستی و سامانهشناسی، منجر به پیدایش ریاضیاتی پیچیده و مدلهای سببی از فرگشت شدند. دیرینهشناسی و کالبدشناسی تطبیقی به ساخت تاریخ فرگشتی حیات با جزئیات بالا کمک کردند. با برآمدن ژنتیک مولکولی در دهه ۱۹۵۰، مبحث فرگشت ملکولی بر پایه رشتههای پروتئینی و تستهای ایمنیشناختی پیدایش یافت، و بعدها بررسیهای بر دیانای و آرانای را دربر گرفت. دیدگاه ژن محور فرگشت اهمیت فراوانی در دهه ۱۹۶۰ یافت، و در پی آن بحثهای زیادی بر روی سازگارپذیری، واحدهای انتخاب، و اهمیت نسبی رانش ژن در مقابل انتخاب طبیعی، انجام گرفتند. در پایان سده ۲۰ میلادی، توالیشناسی دیانای منجر به شناخت فیلوژنتیک مولکولی و انتقال افقی ژن شد، و راه را برای بازنویسی دوباره درخت حیات به صورت سامانهای سه دامنه باز کرد. همچنین، عوامل تازه شناخته شده درونهمزیستی و انتقال عمودی ژن پیچیدگیهای بیشتری را وارد نگره فرگشتی کردند. یافتههای زیستشناسی فرگشتی نه تنها بر شاخههای سنتی زیستشناسی تأثیری بزرگ داشتهاند، که دیگر مباحث علمی و دانشگاهی را نیز (همچون انسانشناسی و روانشناسی) متأثر ساختند.[4]
دایکینگ مولکولی
هنگامی که یک مولکول برای ایجاد یک ترکیب (complex) به مولکول دیگری متصل شود، روش داکینگ مولکولی می تواند جهت گیری مناسب این مولکول در این ترکیب و همچنین شدت پیوند بین این دو مولکول را پیش بینی کند.
هدف از داکینگ پروتئین-لیگاند، پیش بینی جایگاه اتصال مناسب لیگاند به پروتئینی با ساختار سه بعدی شناخته شده است. روش های موفقیت آمیز داکینگ فضاهای با ابعاد بالا را به طور موثر بررسی میکند و از تابع امتیازدهی استفاده می کند تا به درستی کاندیداهای داکینگ را امتیاز بندی کند.
داکینگ مولکولی کاربردهای وسیعی دارد، به طور مثال این روش می تواند برای غربالگری مجازی (virtual screening) كتابخانه (library) های بزرگ ترکیبات، امتیاز دهی به نتایج، و پیشنهاد فرضیه های ساختاری در مورد چگونگی اتصال لیگاند (ligand) به مولكول هدف (target) که در بهینه سازی کاندیداهای دارویی بسيار ارزشمند است و همچنین مطالعات مکانیسم شیمیایی مورد استفاده قرار گیرد.
در مرحله ی ابتدایی، تهیه و آماده سازی ساختارهای ورودی داکینگ به اندازه ی خود داکینگ مهم است. معمولاً ساختارهای اولیه ی مولکول های مورد مطالعه در داکینگ با استفاده از روش های بیوفیزیکی همچون بلورشناسی پرتو ایکس (X-ray crystallography)، طیف بینی رزونانس مغناطیسی هسته (NMR spectroscopy) و میکروسکوپ الکترونی کرایو (cryo-EM) به دست می آیند. البته در مواردی ممکن است این ساختارها با استفاده از روش های مدلسازی مانند همولوژی مدلینگ به دست آمده باشند.
حوزه ی داکینگ مولکولی در دهه های اخیر به واسطه ی نیاز به زیست شناسی مولکولی ساختاری و داروپژوهی (Drug Discovery) برپایه ی ساختار شکل گرفت. داکینگ به واسطه ی پیشرفت و رشد سریع کاربرد و توان کامپیوترها و نیز دسترسی به پایگاه های داده ی پروتئینی و همچنین مولکول های کوچک بسیار آسان شده است. هدف نرم افزار داکینگ مولکولی تشخیص ساختار (molecular recognition) و پیدا کردن جایگاه های اتصال مشابه و پیش بینی شدت اتصال با توجه به انرژی است.
داکینگ مولکولی معمولاً بین یک مولکول کوچک و یک ماکرومولکول هدف صورت می گیرد که در بسیاری از موارد این هدف پروتئین است و این روش تحت عنوان داکینگ پروتئین-لیگاند نامیده می شود. البته توجه به داکینگ پروتئین-پروتئین در حال افزایش است. ماکرومولکول هدف می تواند پروتئین، DNA، و يا RNA باشد.

جمع بندی
تحقیقات کنونی در زیستشناسی تکاملی موضوعات متنوعی را در بر میگیرد و ایدههایی از مناطق مختلف مانند ژنتیک مولکولی و علوم رایانه را در خود گنجانده است.
برخی از زمینههای تحقیق تکاملی سعی در توضیح پدیدههایی دارند که در سنتز تکاملی مدرن به خوبی مورد توجه قرار نگرفتهاند. اینها شامل گونهزایی، تکامل تولید مثل جنسی، تکامل همکاری (فرگشت)، تکامل پیرش و تحول پذیری (فرگشت پذیری) است.
زیست شناسان سادهترین سیر تکاملی را میپرسند: «چه اتفاقی افتاده و چه زمانی؟». این شامل زمینههایی مانند paleobiology و همچنین سامانهشناسی و فیلوژنتیک یا تبارزایش است.
سنتز تکاملی مدرن در زمانی ابداع شد که کسی اساس مولکولی ژن ها را درک نکرد. امروزه زیست شناسان تکاملی سعی در تعیین معماری ژنتیکی پدیدههای جالب تکاملی مانند سازگاری و شکلگری دارند. آن ها به دنبال پاسخ به سؤالاتی از جمله چند ژن درگیر هستند، اثرات هر ژن چقدر زیاد است، تأثیر ژن های مختلف به چه میزانی به هم وابستگی متقابل دارد، تأثیر هر ژن چقدر زیاد است، ژنها چه کاری انجام میدهند و چه تغییراتی در آن ها رخ میدهد (مثلاً جهشهای نقطه در مقابل تکثیر ژن یا حتی تکثیر ژنوم). آنها سعی میکنند با واسطه بررسیهای ارتباطی در کل ژنوم ، وراثت پذیری بالایی را که در مطالعات دوقلوی مشاهده میشود، با این مشکل در پیدا کردن اینکه ژنها در نتیجه این وراثت پذیری تأثیر دارند، تطبیق دهند.
یک چالش در مطالعه معماری ژنتیکی این است که ژنتیک جمعیت کلاسیک که سنتز تکاملی مدرن را کاتالیز میکند باید به روز شود تا دانش مولکولی مدرن را در نظر بگیرد. این امر به پیشرفت زیادی در زمینه ریاضیات نیاز دارد تا بتواند دادههای توالی DNA را با نظریه تکاملی به عنوان بخشی از یک تئوری تکامل مولکولی مرتبط کند. به عنوان مثال، زیست شناسان سعی میکنند با تشخیص رفت و برگشتهای انتخابی، یک ژن را انتخاب کنند.
سنتز تکاملی مدرن شامل این توافق است که نیروها در تکامل نقش دارند، اما در مورد نسبت اهمیت آن ها توافقی ندارند. پژوهش حاضر به دنبال تعیین این موضوع است. نیروهای تکاملی شامل انتخاب طبیعی، انتخاب جنسی، رانش ژنتیکی، پیش نویس ژنتیکی، محدودیتهای رشد، تعصب جهش و جغرافیای زیستی هستند.
یک رویکرد تکاملی برای تحقیقات فعلی در زمینه زیستشناسی ارگانیسم و زیستمحیطی مانند نظریه تاریخ زندگی مهم است. حاشیه نویسی ژن ها و عملکرد آن ها به روش های تطبیقی وابسته است. زمینه زیستشناسی تکاملی تکاملی (“evo-devo”) به بررسی چگونگی عملکرد فرآیندهای توسعه میپردازد و آنها را در ارگانیسمهای مختلف برای تعیین چگونگی تکامل آن ها مقایسه میکند.

- .Mice given bat, provided by University of Colorado Anschutz Medical CampusCold، Spring Harbor Laboratory، University of Bristol.
- https://www.sciencedaily.com/releases/2008/01/080114173923.htm
- https://www.sciencedaily.com/releases/2008/09/080904102756.htm
- https://www.sciencedaily.com/releases/2017/10/171023123815.htm
- SlideShare, https://www.slideshare.net/search/slideshow?searchfrom=header&q=molecular+evolution
- https://www.britannica.com/science/evolution-scientific-theory/The-science-of-evolution#ref49851
- https://www.yourgenome.org/facts/what-is-evolution
- Futuyma, Douglas J., ed. (1999). “Evolution, Science, and Society: Evolutionary Biology and the National Research Agenda” (PDF) (Executive summary). New Brunswick, NJ: Office of University Publications, Rutgers, The State University of New Jersey. OCLC 43422991. Archived from the original (PDF) on 2012-01-31. Retrieved 2014-10-24.
- Futuyma, Douglas J.; Meagher, Thomas R., eds. (2001). “Evolution, Science and Society: Evolutionary Biology and the National Research Agenda”. California Journal of Science Education. Sacramento, CA: California Science Teachers Association. 1 (2): 19–32. ISSN 1531-2488. OCLC 425607451. Retrieved 2014-10-24
- https://ibp.ir،کتاب بیوانفورماتیک سلولی و مولکولی تالیف بابک باباعباسی
- Quanta Magazine ,SlidePlayer
- Lengauer, Thomas, and Matthias Rarey. “Computational methods for biomolecular docking.” Current opinion in structural biology 6, no. 3 (1996): 402-406.Kukol, Andreas, ed. Molecular modeling of proteins. Vol. 443. Totowa, NJ:: Humana Press, 2008.
- https://emri.tums.ac.ir/Modeling/Item/15793 https://cell.ijbio.ir/?_action=article&keywords=%D8%AA%DA%A9%D8%A7%D9%85%D9%84+%D9%85%D9%88%D9%84%DA%A9%D9%88%D9%84%DB%8C&page=2&max_rows=25
- https://rap.sanru.ac.ir/article-1-649-fa.pdf
